


Vol. XXXII Issue 2
Article 6
DOI: 10.35407/bag.2021.32.02.06
RESEARCH
Magrario:
un nuevo genotipo para producir carne ovina de calidad
Magrario: a new genotype to produce quality sheep meat
L.A. Picardi1 *
1 Cátedra de Genética,
CIUNR-IICAR (UNR-CONICET), Facultad Ciencias Agrarias, Universidad Nacional de
Rosario, Campo Villarino, Zavalla (S2125) Pcia. De Santa Fe
* Corresponding
author:
Liliana Picardi lpicardi@unr.edu.ar ORCID 0000-0003-3500-6261
General Editor: Elsa Camadro
RESUMEN
En la producción de carne ovina en
Argentina existe escasa tradición para la comercialización de reses de corderos
pesados con bajo tenor de grasa. Un nuevo genotipo cuya marca registrada es
Magrario (M) fue obtenido en el Campo Villarino de la Facultad de Ciencias
Agrarias de la Universidad Nacional de Rosario (33º S, 61º O) por cruzas
absorbentes de una raza tradicional en la zona, Ideal (I) (Polwarth), hacia la
raza Texel (raza reconocida por producir reses magras). A partir de la
población base de la tercera retrocruza se seleccionó a los machos por el
Aumento Medio Diario relativo hasta los tres meses de edad y a las hembras por
la fertilidad de sus madres. Se verificaron diferencias significativas en la
composición de la res en lo referente a depósitos grasos de este nuevo genotipo
respecto a la población fundadora I tanto en confinamiento posdestete durante
dos meses como en condiciones de cría a campo. También se compararon en
confinamiento corderos M con los de la raza Hampshire Down (HD) siendo los
depósitos grasos significativamente superiores en esta última raza. También se analizaron
cruzamientos de M con HD e I verificando un efecto semi-dominante del nuevo
genotipo para reducir tenor de grasa en las cruzas. La reducción de grasa
subcutánea en el Longissimus Dorsi de (MxHD) se redujo un 20%. Esta experiencia
demostró que Magrario puede ser utilizado como progenitor en cruzamientos si se
desea producir corderos con menores depósitos grasos en condiciones de
suplementación posdestete.
Palabras clave: Ovinos, Carne magra, Confinamiento, Cruzamientos
ABSTRACT
Generally there is poor tradition to produce and to commercialize heavy
lean lamb carcasses. To achieve a better product for the ovine meat market
Ideal (Polwarth) breed ewes were backcrossed to Texel breed rams (breed
recognized to reduce carcass fat). Ideal breed (I) is one of the most ordinary
breeds in Argentina. However, when their lambs are reared in feedlot conditions,
a high fat content is found in their lamb carcasses. After three generations of
backcrosses followed by a breeding program for increase male lamb weaning
weight and female fertility a new genotype was obtained for the local ovine
meat production systems. This new genotype registered as Magrario (M) was
obtained at Villarino Field Station of UNR (Zavalla, Santa Fe, 33º S, 61º W).
It was verify that M produced more lean meat than I breed under feet lot
conditions. M rams were introduced in flocks of Hampshire Down (HD) breed to
evaluate lamb crosses with lean meat. Genotype M was compared under feed lot
conditions with HD lambs during two months in the post weaning. Also crosses
(MxHD), (MxI) and (IxHD) were evaluated in the same conditions. At the end of
the experiment ultrasonic methods were used to evaluate fat depot on
Longissimus dorsi. The (MxHD) showed a reduction of 20% respecting to HD. These
results suggested that M genotype could be a useful paternal genotype to reduce
fat depots when the aim is to produce lamb crosses under feed lot conditions in
a short period of time.
Key words: Ovine,
Lean meat, Feed lot, Crossing
Received: 05/02/2021
Accepted: 05/20/2021
ORIGEN DE LOS OVINOS
Los
ovinos pertenecen a la familia Bovidae, subfamilia Caprinae y al género Ovis,
siendo los ovinos salvajes el muflón (Ovis musimon), el urial (Ovis vignei) y
el argali (Ovis ammon) los ancestros de los ovinos domésticos. Existen
actualmente dos poblaciones de muflones salvajes: el muflón asiático (Ovis orientalis)
y el muflón europeo (Ovis musimon) que frecuentemente en cautiverio pueden
reproducirse entre ellos e incluso pueden dar crías fértiles con ovinos
domésticos (Bunch y Foote, 1977) (Figura 1). Para algunos
autores (Zarazaga et al., 1978) la taxonomía de los miembros del género Ovis
estaba sujeta a controversias. Sin embargo hay trabajos más recientes (Chessa
et al., 2009) que con nuevas metodologías aportan al conocimiento de la posible
genealogía de Ovis. Todas las razas de ovinos domésticos tienen un cariotipo de
2n=54, idéntico al del muflón europeo (Ovis musimon), el muflón asiático (Ovis
orientalis) y al de los ovinos norteamericanos Bighorn (Ovis canadienses) y
Dall (Ovis dalli). Junto con la cabra, el ovino ha sido la especie que fue
utilizada por el hombre en forma temprana en su camino a la producción de
alimentos y abrigo. Sus parientes salvajes tenían algunas características que
las hacían especialmente adecuadas para la domesticación, tal como su
relativamente poca agresividad, un tamaño manejable, una pronta madurez sexual,
su carácter gregario y su alta tasa de reproducción. Si bien hay evidencias por
rastros arqueológicos que la domesticación de ovinos en la Mesopotamia fue alrededor
de 9000 años A.C., es recién en 3000 A.C. que se encontraron evidencias en
tablas de arcilla de representaciones artísticas y escritos sobre los ovinos.
En tiempos de los babilonios y los asirios el ovino tenía lana y una cola larga
grasa, lo que permite suponer que así almacenaban energía para sobrevivir en
zonas áridas. También hay registros con majadas de cientos de ovinos para carne
con fenotipos totalmente distintos, lo que evidenciaría la importancia de esta
especie para la economía de esta sociedad.

Figura 1. Muflón europeo (izq.) y Merino actual
(der.)
Se
considera que los primeros ovinos que llegaron a suelo rioplatense fueron
traídos por Nuflo de Chaves, que en 1549 los llevó desde Lima (Perú) a Asunción
(Paraguay) (Giberti, 1981). Para los conquistadores los vacunos y los ovinos
eran necesarios para la provisión de carne y otros productos, siendo los ovinos
los más fáciles de desplazar y es así que en el Litoral y en el NOA los ovinos
se difundieron antes que los vacunos. A esto se puede sumar una mayor
adaptación a las condiciones ambientales, con menores exigencias de pastoreo,
mansedumbre y la asistencia de los pobladores indígenas para su atención y
esquila. Estos ovinos recién llegados al nuevo mundo serían ovejas de razas
sirias, pirenaica y berberisca, ya que la corona de España tenía prohibido la
exportación de las finas ovejas Merino (Wernicke, 1933). De hecho las ovejas
criollas que se encuentran en el norte de nuestro país tienen un mayor parecido
fenotípico con la raza Churra española que con la Merino (AACM, 2000).
LA HISTORIA DEL OVINO EN NUESTRO PAÍS
Se
ha registrado que para 1810 nuestro país tenía alrededor de tres millones de
ovinos de baja calidad que denominaban como raza Criolla, con cuerpo pequeño,
lana de diferentes colores, escasa, corta y enrulada. En 1813, a instancias del
cónsul de Estados Unidos, llegó el primer plantel compuesto por 100 ovejas
Merino con sus respectivos carneros, dando origen a la primera cabaña argentina
de ovinos, ubicada en el actual partido de Morón (Pcia. Buenos Aires). En 1824,
durante el gobierno de Rivadavia, fueron introducidos 100 Merinos más de España
y 30 animales de la raza Southdown de Inglaterra con destino a la futura cabaña
“Los Galpones” en las vecindades de San Vicente. A fines de 1826 Rivadavia
importa otro lote de 150 Merinos finos que son comprados por los mismos dueños
de “Los Galpones”, cabaña que para 1830 gozaba de enorme prestigio por los
resultados de sus ventas, circunstancia que generó un auténtico interés por el
Merino o “merinomanía”. Como el Merino español no podía salir de la península,
se importaban ejemplares de Sajonia, descendientes de los “Negretes” españoles.
A través de sucesivas importaciones, entre 1836 y 1838, ingresan al país un
total de 7850 animales. Se destacaron por entonces como criadores de ovejas,
los irlandeses, ingleses y escoceses, expertos en este tipo de actividad por
haberla ejercido en su país natal. Muchos de esos extranjeros compraron campos
a precios irrisorios a Unitarios que perseguidos por Rosas, los vendían antes
de que éste los embargase. Estas persecuciones políticas hicieron perder la
afición por el lanar, sin embargo ante la caída de Rosas y la posterior
normalización de las corrientes comerciales se retomó el proceso de desarrollo
lanar que se había iniciado con anterioridad. Por los años 1850-55, dado los
logros económicos obtenidos con los ovinos por estancieros del norte de la
provincia de Buenos Aires, acompañado por el interés de la industria europea
para manufacturar tejidos que requerían lana larga, se produjo un mayor interés
por la cría de ovinos. Es así que durante los siguientes cuarenta años la lana
ocupó el primer lugar entre las exportaciones argentinas y el tercer lugar en
el mundo como país productorexportador de lanas. Posteriormente, cambios
operados en la estructura agraria de la provincia de Buenos Aires, condujeron a
la declinación de la cría del lanar y a su desplazamiento hacia zonas
extra-pampeanas como la Patagonia, que para 1888 tenía apenas 300.000 lanares.
Actualmente en la Argentina según SENASA (2000) se estiman alrededor de
14.000.000 de cabezas. La mayor proporción se encuentra en la Patagonia donde
las bases genotípicas han sido las razas Corriedale y Merino, si bien con el
tiempo se han sumado otras razas terminales, cruzas o biotipos que han emergido
de estas razas. Estas poblaciones se han adaptado a condiciones de crías
extensivas en el medio ambiental patagónico. Las Asociaciones de Criadores de
Merino y Corriedale, entre otras, han colaborado en el programa de Cordero
Patagónico que es altamente reconocido en el país y el exterior. Asimismo las
Asociaciones de Criadores Ovinos participan con el INTA en programas de gran
impacto para el país como son el PROVINO y PROLANA.
Después
de la Patagonia le siguen en importancia para la producción ovina la región
pampeana, el litoral y por último la región del NOA. Los sistemas productivos
varían de norte a sur, con objetivos de subsistencia, lanero, carnicero,
lechero o mixtos. Por lo tanto según el objetivo productivo y la región se
utilizan distintas razas y por eso en esta especie existen múltiples razas y
ecotipos adaptados a los más variados ambientes. Se puede calificar a las razas
actuales en genotipos apropiados para la producción de carne y/o producción de
lana. Las razas que han sido seleccionadas para calidad de lana son
descendientes de la raza Merino. Hay otras razas que tienen doble propósito,
lana y carne y que fueron introducidas al país en distintas épocas tal como la
citada Corriedale, las Lincoln, Romney Marsh y la raza Hampshire Down, que es
netamente productora de carne y tiene presencia en la región pampeana y el
litoral (Calvo, 1982). Otra raza reconocida en nuestro país por su calidad de
lana es la Polwarth. Su origen se remonta a 1880 en Australia (Victoria,
condado de Polwarth) por el cruzamiento de la Merino con la raza Lincoln para
mejorar largo de mecha y mantener la finura siendo su proporción según las
razas progenitoras: ¾ Merino y ¼ Lincoln. Esta raza, que fue introducida en
Uruguay alrededor de 1938, adquiere el nombre de raza Ideal por sus criadores
dada su excelente calidad de lana y rusticidad. Posteriormente se introdujo en
nuestro país, principalmente en establecimientos laneros del Litoral y la Pampa
Húmeda (Figura
2).

Figura 2. Raza Ideal (Polwarth)
En
los últimos años se ha introducido en el país la raza Texel, reconocida como
raza esencialmente productora de carne de calidad. Esta raza proviene de un
grupo de ovejas de cola corta que tiene una marcada distancia genética de la
Merino (Gines, 2007) y se originó en la isla Texel situada en el noroeste de
Holanda. Al final del siglo XIX un reducido núcleo de esta raza fue cruzada con
las razas inglesas Lincoln y Leicester con el fin de obtener un tipo de animal
de bajo tenor de grasa en la carcasa. Actualmente se la reconoce por esta
cualidad y se ha difundido en Europa como raza terminal para producir carcasas
de carne magra (Leymaster y Jenkins, 1993; Visscher, 2000). Al continente
americano llegó con su introducción en Uruguay hace más de 40 años y desde ahí
llega a nuestro país introduciéndose como raza productora de carne en la Pampa
Húmeda y el alto valle del Río Negro (Figura 3).

Figura 3. Raza Texel
OBTENCIÓN DEL NUEVO GENOTIPO MAGRARIO
En
nuestro país la producción de carne ovina se basa esencialmente en la oferta
del cordero “lechal” o sea al destete. Respecto a la producción de carne ovina
existe escasa tradición en la comercialización de corderos pesados con bajo
tenor de grasa en la res (Bianchi et al., 2005). Las poblaciones ovinas en el
sur de la Pcia. de Santa Fe constituyen generalmente un mosaico racial, y
cuando los corderos se suplementan para producir corderos pesados las reses
suelen depositar un nivel de grasa no aceptado por el consumidor. Este
consumidor, influenciado por el actual sistema social, está dedicado
actualmente a procurarse alimentos magros para su dieta, hecho que incluso se
observa en la Comunidad Europea (Bermués et al., 2012; Higgs, 2000; Fogarty,
2009). La producción de corderos pesados con escasos depósitos grasos
resultaría una alternativa para los pequeños productores ovinos si cuentan con
genotipos adecuados que no depositen grasa en la res, aún en condiciones de
confinamiento. Un cambio de objetivo productivo, como es la producción de un
cordero pesado (de 40 kg a 45 kg) implica contar con un tipo de animal que no
deposite grasa ya que como se señaló esta condición no es aceptada en los
consumidores (Higgs, 2000; Hopkins et al., 2008; Bianchi et al., 2005).
Bajo
el objetivo de ofrecer a los productores de la Pcia. de Santa Fe un nuevo
enfoque para producir carne de calidad, se inició 1986 en el Campo Experimental
Villarino de la Fac. Cs. Agrarias de la UNR en Zavalla (33º S, 61º O) un
programa de retrocruzas de la raza Ideal hacia la raza Texel. Se inició un
programa de retrocruzas con una majada 100 madres Ideal (I) hacia la Texel (T)
con tres machos de la Estancia María Luisa (Cnel. Vidal, Pcia. de Bs. As.).
Otras 100 madres Ideal fueron cruzadas con otros tres machos originados en la
Estancia Tres Lanzas (Pcia. Río Negro) con el fin de contar con distintos
aportes genéticos de la raza Texel. Después de tres retrocruzas (BC3) en una
población base de 329 animales se inició un programa de selección donde los
machos al destete (3 meses de edad) se seleccionaron por el AMDr (Aumento Medio
Diario relativo) desde el nacimiento al destete como una medida de eficiencia
de conversión de alimento (AMDr=cociente entre AMD y el peso medio (Pm) en el
período estudiado: (Pm=(PInicial + PFinal)/2) (Fitzhugh y Taylor, 1971; Esteva
y Picardi, 1989). Las hembras se seleccionaban por la fertilidad de sus madres
que fue evaluada a través de un índice (Picardi y Rabasa, 1984; Toso y Picardi,
1995). La endocría se limitó manteniendo un alto número efectivo (Ne) (Falconer
y Mackay, 1996). Después de varios ciclos de este programa este nuevo genotipo
obtuvo la marca registrada de Magrario (M) (Picardi, 1999).
EXPERIMENTOS EN CONFINAMIENTO POSDESTETE
Para
verificar diferencias en la calidad de las reses de este nuevo genotipo con las
de la raza progenitora Ideal en producir corderos pesados se llevó a cabo un
experimento con Magrario e Ideal en confinamiento posdestete (n=68 animales de
ambos sexos) y a campo posdestete(n=99 animales de ambos sexos). Se aplicó la
metodología de la EEAP Standard Methods of Sheep Carcass (Fisher y De Boer,
1994) para evaluar 22 caracteres en las reses después de dos meses en estas
condiciones de cría (Toso et al., 1995; Acebal et al., 1997; Acebal et al.,
2000 ). Se encontraron diferencias significativas en el Peso Final después de
los dos meses en confinamiento entre M e I (M=40,2±0,8 vs. I=36,7±0,9;
p<0,01). Entre las características de la res que mostraban las diferencias
entre estos genotipos se destacaron el peso de la res en frío (M=18,7±0,6 vs.
I=15,7±0,7; p<0,01) y el peso de carne magra total (M=5,5±0,2 vs. I=4,2±0,1;
p<0,001). Fue interesante comprobar que no hubo diferencias significativas
entre las corderas M e I en ninguno de los ambientes, lo cual podría deberse al
dimorfismo logrado por efecto de la selección de la fertilidad femenina en M,
ya que no se consideró la ganancia de peso para este sexo (Toso y Picardi,
1995; Picardi et al., 2006). Bradford (2002) ya había señalado que los
mejoradores debían ser cuidadosos al querer modificar el tamaño en las hembras
pues esto puede afectar la aptitud reproductiva. También se ha señalado que a
veces las diferencias en las reses entre sexos pueden deberse al incremento de
depósitos grasos en las hembras al seleccionar por tamaño. La evaluación de las
reses siguiendo el protocolo de la EEAP y un Análisis Multivariado de
Componentes Principales posterior dentro de cada ambiente, permitieron definir
que los corderos Magrario e Ideal constituyen distintos grupos
independientemente de las condiciones ambientales en las cuales son criados
(Picardi et al., 2010). Asimismo se verificó que este nuevo genotipo ovino
tiene lana blanca con una finura promedio de 26 micras según los análisis del
laboratorio de fibras de INTA Bariloche.
Si
bien esta experiencia ofreció importantes datos sobre la composición de la res
de los Magrario, este tipo de evaluación conlleva al sacrificio de los animales
y por lo tanto no es de utilidad para la clasificación de futuros
reproductores. En consecuencia se aplicó en la majada las técnicas de
ultrasonido para evaluar los depósitos grasos en el Longissimus dorsi ya que es
considerado un buen predictor del tenor de grasa en las reses (Kvame y Vangen,
2007; Lambe et al., 2009). Esta metodología fue usada también en otro
experimento en el Campo Experimental Villarino con el fin de comparar ahora los
corderos Magrario con una raza productora de carne. Entre las razas que se
encuentran en nuestra zona, la Hampshire Down (HD) es la más reconocida para
producir carne (Mueller, 2005).
Si
bien no tiene lana de calidad, sus corderos se han establecido como los de
mayor frecuencia para ofrecer al consumidor carne ovina. Para evaluar depósitos
grasos en la res se utilizó la técnica de ultrasonido con la cual se esquila el
área elegida y se ubica el punto de medición para utilizar el equipo de
ultrasonido por palpación de la 13er costilla, que fue establecido a la altura
de la vértebra dorsal correspondiente, 4 cm hacia lateral de la apófisis
espinosa y 4 cm hacia la craneal de la misma. La metodología de ultrasonido
permitió obtener sobre el L. dorsi las siguientes mediciones: área (AL, cm2);
grasa subcutánea (GS, cm); grasa periférica (GP, cm); y veteado (V,
infiltración grasa intramuscular) (Figura 4). Después de dos
meses en confinamiento posdestete no se obtuvieron diferencias significativas
en el Peso Final (PF, kg); PF (M)=39,3±0,6 y PF (HD)=41,5±1,0 ni en AL (cm2):
AL (M)=20,7±1,3 y AL (HD)=18,0±1,8. Sin embargo, las diferencias fueron
altamente significativas (p<0,001) para GS (cm): GS (M)=0,48±0,02 vs. GS
(HD)=1,27±0,08 y para GP (cm): GP (M)=0,31±0,02 vs. GP (HD)=0,53±0,03. Para V,
característica visual de importancia económica, también hubo diferencias
altamente significativas: V (M)=0,31 y V (HD)=5,2 (Relling et al., 2010) (Figura
5).
Figura 4.
Magrario con 6 meses de edad con evaluación del L. dorsi por ultrasonido

Figura 5. Magrario al finalizar el confinamiento
posdestete de dos meses
Con
estas experiencias en la majada Magrario se pudo comprobar la eficacia de las
incorporaciones de los genes Texel para producir un nuevo genotipo ovino
productor de carne magra. Sin embargo para los productores ovinos no es posible
la incorporación de este genotipo en sus explotaciones en forma total. Por lo
tanto la incorporación de machos Magrario en cruzamientos con sus majadas
podría ser una forma rápida de cambiar el tipo de cordero a producir. Para ello
era necesario evaluar si las cruzas con Magrario presentaban características de
este genotipo en las F1. Con este objetivo se condujo una ensayo de
cruzamientos de hembras I con machos M (IxM, n=23) y HD (IxHD, n=22) y hembras
M con machos HD (MxHD, n=21). Se evaluaron corderos y corderas de estas cruzas
en dos meses de confinamiento posdestete (siendo los testigos los genotipos M y
HD) en las siguientes variables: peso inicial y final y el cálculo individual
de la eficiencia de conversión de alimentos con el AMDr. (Picardi et al., 2017).
Al finalizar este período se utilizó la técnica de ultrasonido para evaluar los
depósitos grasos en el L. dorsi (Hopkins et al., 2008) y se obtuvieron las
siguientes variables: Espesor de Grasa Subcutánea (GS, cm); Grasa Perimuscular
(GP, cm); Dimensión Longissimus Ancho (DLA, cm); y Dimensión Longissimus
Profundidad (DLP, cm).
Con
respecto a las variables de biomasa para M y HD los valores del peso al
finalizar el período fueron similares al igual que los valores de AMDr. En los
valores obtenidos sobre el L. dorsi se encontraron nuevamente diferencias
altamente significativas entre M y HD para GS y GP evidenciando que, si bien
ambos genotipos alcanzan el mismo peso, HD lo hace aumentando su tenor de
grasa. Es de destacar que todos los genotipos fueron igualmente eficientes bajo
estas condiciones ambientales tal como se refleja en la estimación de
eficiencia a través del AMDr. Si bien el peso final que alcanzan los corderos M
resultó similar a los HD, indicando que estos corderos tendrían igual biomasa,
las diferencias estarían en los tenores de grasa detectados por la metodología
del ultrasonido. Según estas observaciones los corderos M resultarían más
eficientes para convertir el alimento en proteína más que en grasa (Relling et
al., 2010).
Cuando
se analizó el grado de heterosis para PF y las evaluaciones de la grasa en L.
dorsi, caracteres que definirían las ventajas productivas de las cruzas aquí
evaluadas, se observó en los corderos machos una reducción del 10,5% para PF
respecto al progenitor HD y de 19,7% para GS, mientras que para GP la reducción
fue del 17% con respecto a los corderos HD. Pero si se realizaba la comparación
de la cruza MxHD, respecto a corderos del genotipo progenitor M, en el PF hubo
una disminución de 11,9% mientras que para GS en MxHD hubo un aumento de 27% y
en GP del 10% (Figuras 6 y 7). Lo más destacado de este experimento
fue que si bien las cruzas no presentan un aumento significativo en la biomasa,
y por ende en los kilos a producir, la utilización de machos M permitiría
obtener corderos cruzas con menores depósitos grasos que la otra raza HD si se
tiene como objetivo obtener corderos pesados magros en confinamiento en corto
plazo. Si bien no hubo grados significativos de heterosis para el peso y la
eficiencia, se debe considerar que la heterosis para ganancia diaria puede incrementar
en la medida que aumenta la edad del animal según Bianchi et al. (2005).

Figura 6. Corderos F1 (MxHD) y Magrario después de
dos meses de confinamiento posdestete

Figura 7. Madre y cordero Magrario (izq.) - Madre
Ideal con cordero (IxM) (der.)
Otro
análisis que permitió ubicar a las cruzas con respecto a los genotipos M y HD
en su rendimiento se obtuvo con un análisis de conglomerados. Este análisis
multivariado, con todos los caracteres evaluados, permitió determinar la
ubicación de las cruzas respecto de los progenitores M y HD en distintos
agrupamientos. En estos dendrogramas pudo observarse que el genotipo M, tanto
en corderos machos como hembras, se diferencia de los restantes genotipos
siendo mayor la distancia observada con corderos machos HD. La conformación de
los grupos que este análisis permitió obtener demostró que los genotipos cruzas
se ubican cercanos a M, especialmente las cruzas donde M es progenitor (Figura
8).
La correlación cofenética fue de 0,85 para machos y 0,80 para hembras; si bien
para este sexo el genotipo más distante de M fue la cruza IxHD y la más cercana
MxHD. La visualización de esta conformación grupal justificaría el supuesto de
que el genotipo Magrario actuaría con una acción génica semidominante en
combinación con los otros genotipos raciales en este estudio.
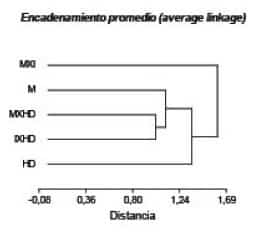
Figura 8. Análisis de conglomerados considerando
todas las variables para corderos machos de los genotipos Magrario (M),
Hampshire Down (HD), la F1 (MxI), F1 (MxHD) y F1 (IxHD).
CONCLUSIONES
Este
proyecto de investigación en genética ovina fue realizado en el Consejo de
Investigaciones de la UNR y ha permitido la obtención de un nuevo genotipo
ovino productor de carne de calidad, pero también tuvo impacto en la zona por
la difusión de un nuevo modelo productivo para los pequeños productores. El
Campo Experimental Villarino fue un módulo productivo para la obtención de
corderos pesados promovidos por la Ley Provincial Ovina Santafesina Nº 12483
(adherida a la Ley Nacional Nº 25422). Bajo este programa de extensión se
desarrolló un protocolo para producir cortes en las reses con el protocolo
neozelandés y su posterior envasado al vacío lo que le da valor agregado a la
producción de carne ovina pudiendo establecer de esta forma una cadena de valor
para distintos actores de la producción. Es así, que al presentar en un Foro
para Latinoamérica estos resultados se destacó que “Planteada en términos de
diversificación productiva, la carne ovina es una alternativa tanto para
productores pequeños que conservan o planifican reflotar sus chacras mixtas. El
aumento de productividad de las majadas asociado a innovaciones de producto y
proceso permitirían generar una oferta continua, adecuada a los requerimientos
del mercado. Implicaría una modernización de las tradiciones productivas ovinas
al incorporar tecnología y organización y una mejora competitiva para la
sostenibilidad de los sistemas agropecuarios” (Picardi et al., 2016).
AGRADECIMIENTOS
Este
trabajo contó con la colaboración en la Universidad Nacional de Rosario de las
Cátedras de Anatomía y Fisiología Animal y de Nutrición Animal de la Facultad
de Ciencias Agrarias, y de la Cátedra de Producción de Porcinos y Pequeños
Rumiantes de la Facultad de Ciencias Veterinarias.
BIBLIOGRAFÍA
AACM
(2000) Asociación Argentina de criadores de Merino: La raza Merino en la
Argentina. Artes Gráficas Corin Luna, Buenos Aires.
Acebal M.A., Maiztegui L.B., Amelong
J., Picardi L.A. (1997)
Evaluación de características de la carcasa en corderos cruza de la raza Ideal
con la Texel en confinamiento y a campo. Archivos Latinoamericanos de
Producción Animal 5 (1): 552-554.
Acebal M.A., Maiztegui L., Amelong
J., Picardi L.A. (2000)
Evaluación de características de la canal en corderos con ¾ de genotipo de la
raza Texel. Archivos Latinoamericanos de Producción Animal 8 (2): 55-58.
Bernués A., Ripoll G., Begoña P.
(2012) Consumer segmentation based on convenience orientation and attitudes
towards quality atributes of lamb meat. Food Quality and Preference 26: 211-220.
Bianchi G., Garibotto G., Bentancur
O., Feed O., Franco J., Peculio A., Sañudo C. (2005) Características productivas y
calidad de carne en corderos pesados Corriedale y Hampshire Down x Corriedale. Rev. Arg. Prod. Animal 25: 75-91.
Bradford, E. (2002) Breeding and
selection- In Sheep Production Handbook- American Sheep Industry Association, Fort
Collins, USA
Bunch T.D., Foote W.C. (1977) Evolution
of the 2n=54 karyotype of Domestic sheep (Ovis aries). Ann. Génét. Sél. anim.
9 (4): 509-515.
Calvo C. (1982) Ovinos. Orientación Gráfica
Editora S.R.L. Buenos Aires, Argentina.
Chessa F., Pereira F., Arnaud A.,
Amorim F., Goyache I., Mainland R.R., Kao J.M., Pemberton D., Beraldi M., Stear
A., Alberti M., Pittau L., Iannuzzi M.H., Banabazi R., Kazwala Y.P., Zhang J.J.,
Arranz B.A., Ali Z., Wang M., Uzun M., Dione I., Olsaker L.E., Holm U., Saarma S.,
Ahmad N., Marzanov E., Eythorsdottir M.J., Holland P., Ajmone Marsan M.W., Bruford
J., Kantanen T.E., Spencer T.E., Palmarini M. (2009) Revealing the history of
sheep domestication using retrovirus integrations. Science 324 (5926): 532-536.
doi:10.1126/science.1170587.
Esteva J., Picardi L.A. (1989) Eficiencia
postdestete en corderos de la raza ideal y sus cruzas y retrocruzas con la raza
Texel. Rer. Arg. Prod. Animal 9 (6): 457-462.
Falconer, D.S. and Mackay, T.F.C.
(1996) Introduction to Quantitative Genetics. -4th Edition. Longman Group Ltd.
Fisher A.V., de Boer H. (1994) The
EEAP Standard Methods of sheep carcass assessment, carcass measurements and
dissection procedures. Livestock Production Science 38: 149-159.
Fitzhugh H.A., Taylor ST.C.S. (1971)
Genetics analysis of degree of maturity. J. Anim. Sci. 33: 717-725.
Fogarty N.M. (2009) Meat sheep
breeding - Where we are at and future challenges. Proc. Assoc. Ad.
Anim. Breed Genet. 18: 414-421.
Giberti H.C. (1981) Historia
económica de la ganadería en la Argentina. Ed. Solar, Argentina.
Ginés S. de GEA (2007) El ganado
lanar en la Argentina. Ed. NRC, 2da Ed., Argentina.
Higgs J.D. (2000) The change
nature of read meat. Trends Food Sci. Technol. 11: 85- 95.
Hopkins D.L., Ponnampalam E.N., Warner
R.D. (2008) Predicting the composition of lamb carcasses using alternative fat
and muscle depth measures. Meat Sience 78: 400-45.
Kvame T., Vangen O. (2007) Selection
for lean weight based on ultrasound and CT in a meat line of sheep. Livestock Science Vol. 106 (2-3): 232-242.
Lambe N.R., Navajas E.A., Fisher A.V.,
Simm G., Roehe R., Bünger L. (2009) Prediction of lamb meat eating quality in
two divergent breeds using various live animal and carcass measurements. Meat Science Vol. 83 (3): 366-375.
Leymaster K.A., Jenkins T.G. (1993)
Comparison of Texel-sired and Suffolk-sired crossbred lambs for survival,
growth and compositional traits. J. Anim. Sci .71: 859-86.
Mueller J. (2005) Síntesis de
las razas ovinas y su uso en la Argentina. Memorias del VII Curso de
Actualización en Producción Ovina, EEA Bariloche, INTA
, Argentina.
Picardi L.A. (1999) Marca
Registrada MAGRARIO- Acta Nº 2222.703(51) Clase 29. Registro de Propiedad,
Instituto Nacional de Propiedad Industrial (Secretaría de Industria y Comercio
República Argentina).
Picardi L.A., Acebal M., Maiztegui
L. (2006) A new ovine genotype to improve lamb meat quality. 8th World Congress
of Genetics Applied to Livestock Production, Belo Horizonte, Brasil.
Picardi L.A., Keilty H., Maiztegui
L.B., Amelong J., Acebal M.A., Piga E. (2017) Evaluación del nuevo genotipo Magrario
como progenitor en cruzamientos con otras razas ovinas. Rev. Arg. Prod. Animal Vol. 37: 33-39.
Picardi L.A., Maiztegui L., Acebal
M. (2010) Verifying carcass traits in a backcross programme with Texel Breed. Livestock
Science 127:
267-271.
Picardi L.A., Principi L., Keilty
H. (2016)
Carne de calidad como instrumento de gestión tecnológica para un desarrollo
sostenible en el Sur de la Pcia. de Santa Fe, República Argentina. II Foro Regional
de Innovación para el desarrollo sostenible, Santiago, Chile.
Picardi L.A., Rabasa S.L. (1984) Efecto de la
selección divergente de peso sobre los parámetros de la curva de crecimiento y
la eficiencia de conversión en ratones. Mendeliana VI (2): 43-47.
Relling A.E., Gaeta N., Pellejero
L., Keilty H., Picardi L. (2010) Deposición de grasa en dos genotipos ovinos
durante el posdestete. Rev. Arg. Prod. Animal Vol.
30 (1): 433-434.
Toso A., Acebal M., Calvo F., Picardi
L.A. (1995)
Crecimiento posdestete en confinamiento y a campo de corderos de la raza Ideal
y su retrocruza hacia Texel. Rev. Arg. Prod. Animal 5 (3/4): 936-939.
Toso A., Picardi L.A. (1995) Relación entre
la tasa de madurez y la fertilidad en ovinos. Actas 1era Jornada de Genética
Argentino Chilena, XXVI Congreso Argentino de Genética. Bariloche, Argentina; p. 90.
Visscher A.H. (2000) The
influence of the Texel breed on European sheep production. Book of Abstracts
EAAP (6), 51st Annual Meeting of EEAP, Wageningen. The Netherlands.
Wernicke E. (1933) El paso del
ganado lanar desde el antiguo al nuevo mundo. Anales de la Sociedad Rural
Argentina N° 7.
Zarazaga I., Arruga V., Vallejos M.
(1978) Livestock Cytogenetics. I. Ovis aries. An. Aula Dei. 14 (1/2): 128-140.